해조류 유래 황산화 다당류; 뉴에이지 화학예방제: 종합적인 검토
Sep 22, 2023
간단한 요약: 황산화다당류는 강력한 화학치료제 또는 화학예방제로서 여러 암세포주에서 면역력을 높이고 세포사멸을 촉진하여 항암 특성을 가지고 있습니다. 황산화 다당류는 상당한 항산화 및 면역 조절 잠재력을 갖고 있으며, 이는 낮은 세포 독성으로 질병 예방 효과에 기여하고 동적 세포사멸 조절을 통해 암에 대한 우수한 효능 치료 결과를 제공합니다. 또한 식이 보충제나 암에 대한 보조 치료제로도 사용할 수 있습니다.

cistanche tubeulosa-항종양의 장점
추상적인: 해조류 유래 생리활성 화합물은 인간의 질병 치료에 정기적으로 사용됩니다. 황산화 다당류는 발견된 이후로 강력한 화학요법 또는 화학 예방 약물입니다. 그들은 면역력을 강화하고 세포사멸을 촉진함으로써 항암 특성을 나타냈습니다. ROS 생성 제어, 필수 세포 생존 및 사멸 과정 보존과 같은 중요한 세포 내 신호 전달 경로의 동적 조절을 통해 황산화 다당류의 항산화 및 면역 조절 잠재력은 질병 예방 효과에 기여합니다. 황산화 다당류는 암에서 세포사멸의 동적 조절을 통해 낮은 세포 독성과 우수한 효능의 치료 결과를 제공합니다. 황산화 다당류가 인간 암세포에 어떻게 영향을 미치고 세포 사멸 경로에 분자가 관여하는지 이해하면 새로운 화학 예방 방법이 제시될 것입니다. 이 리뷰에서는 세포사멸과 자가포식 조절 황산화 다당류의 중요성뿐만 아니라 더 큰 임상 효능을 위한 향상된 나노제형의 미래 방향이 강조되었습니다. 더욱이, 이 리뷰는 황산화 다당류의 화학요법적 사용의 가능한 메커니즘, 항암제로서의 잠재력, 다양한 악성 종양에서 세포사멸을 유도하기 위해 제안된 작용 메커니즘에 대한 최근 연구 결과에 초점을 맞추고 있습니다. 독특한 물리화학적 및 생물학적 특성으로 인해 황산화 다당류는 질병에 대한 기능과 적용을 향상시킬 수 있는 생리 활성 성분으로 이상적입니다. 그러나 황산화 다당류의 물리화학적 특성 및 기능성과 기능성 암에서의 황산화 다당류 기반 전달 시스템의 사용에 관한 문헌에는 공백이 있습니다. 또한, 전임상 및 임상시험을 통해 암에 대한 약물의 효능이 밝혀질 예정이다.
키워드: 세포사멸; 암; 화학적 예방; 황산화 다당류
1. 소개
현재 전 세계적으로 인구가 폭발적으로 증가하고 식습관 및 생활 방식의 변화가 질병 발생의 중요한 요인으로 간주됩니다. 알츠하이머병, 파킨슨병, 당뇨병, 암 및 기타 신경 질환과 함께 수많은 감염으로 인한 질병이 인간 수명에 심각한 위험을 초래합니다[1]. 확인되지 않은 세포 성장으로 인해 발생하는 수많은 병리학적 문제의 집합체인 암은 개인의 건강 관리에 해로운 영향을 미칩니다[2]. 2019년 추정에 따르면 전 세계적으로 매년 960만 명 이상의 사망을 초래하는 치명적인 암 종류는 200종 이상입니다[3]. 주요 사망원인은 피부암, 위암, 유방암, 폐암, 전립선암, 대장암 등이다[4]. 저소득 및 중간 소득 국가의 사망률 사례는 약 70%로 기록되어 있습니다[5]. 역학 연구를 바탕으로 세계보건기구(WHO)는 2018년 암 관련 사망자가 960만 명, 신규 암 환자가 1,800만 명에 달할 것으로 예측했습니다[6]. 여러 항상성 시스템은 암세포의 통제되지 않은 증식으로 인해 파괴되어 유전적 변화로 인해 암세포의 침입과 전이를 초래합니다[7]. 암 치료 방법에는 수술, 화학요법, 방사선요법, 면역요법 등이 있으며; 화학 요법이 가장 자주 사용됩니다 [7-9]. 화학요법은 암세포와 비암세포 모두에서 세포독성을 유발하여 여러 중요한 기관을 손상시키는 일반적이고 효율적인 암 치료법입니다[10]. 암 치료에서는 부작용과 심각한 반응을 제거하기 위해 약물 내성이 주요 관심사이다[11,12]. 따라서 부작용이 적고 내약성이 높은 항암제를 개발하고 찾는 것이 중요하다. 화학요법은 때때로 표적 부위 주변에 불리한 환경과 돌이킬 수 없는 장기 손상을 초래합니다. 추가적으로, 약물에 대한 세포적 관대함은 추가적인 치료적 어려움을 야기합니다. 따라서 모든 불리한 조건을 견딜 수 있는 부작용이 적은 새로운 치료제를 찾는 것이 요구된다[13]. 약물로 사용되는 자연 발생 생체 활성 화합물은 다양한 치료 용도를 가지고 있습니다 [14-18]. 또한 대부분의 의약품은 천연 화합물이거나 합성 등가물입니다 [19]. 다양한 화학적 구성과 생물학적 이용 가능성으로 인해 해양 천연물(MNP)이 최근 치료 후보로서의 잠재력에 대해 조사되었습니다[20-25]. 조류 생물다양성은 해양 및 담수 환경에 풍부하며 주요 생리활성 대사산물에 기여합니다[24,26-36]. 해조류는 민물과 바닷물 모두에서 발견되며 해양 생태계의 생태와 생물 다양성을 보존하는 데 중요한 역할을 합니다[37,38]. 황산화 다당류의 항응고제, 항암제, 항당뇨병제, 항바이러스제, 면역조절제, 항혈관신생제, 항염증제, 항부착제 및 항신경퇴행성 특성은 잠재적인 치료제로 사용됩니다[39-44]. 후코이단, 포르피린, 카라기난, 울반은 갈조류, 홍조류, 녹조류에서 흔히 추출되는 황산화 다당류이며 많은 악성 종양에 대한 치료적 근면성이 있는 황산기를 함유하고 있습니다[45-47]. 황산화 다당류는 엄청난 구조적 다양성과 강력한 항산화 능력으로 인해 임상에서 화학요법 약리제로 사용될 것으로 예상됩니다 [48]. 또한 높은 흡수력, 저렴한 유지 비용, 향상된 생산 및 식품 보충제로 사용하면 더욱 인기 있는 화학 요법 약물이 됩니다. 이전 보고서에서는 조류 유래 황산화 다당류의 항암 특성에 대해 논의했습니다[50-52]. 그러나 정확한 메커니즘은 완전히 이해되거나 논의되지 않습니다. 더욱이 상황별 약물 표적, 암 하위 유형 및 종양 미세 환경은 논의되지 않습니다 [52-54]. 원고의 현재 형태는 세포 사멸 경로 유도에 이러한 조류 유래 황산화 다당류의 기계적 관여를 논의합니다. 종양의 이질성을 염두에 두고 다중 표적 특정 단일 약물 요법도 논의되었습니다. 암 예방에 사용되는 황산화 다당류 조절 경로의 기원에 대한 체계적인 분석과 기계적 개요가 이 검토의 주요 주제입니다. 암 예방의 맥락에서 치료적 개입을 이해하기 위해 우리는 화학적 복잡성과 황산화 다당류의 공급원에도 집중했습니다. 이 검토에서는 미래의 임상 및 나노 약물 전달 용도를 제안하며, 이는 암 예방에서 황산화 다당류의 잠재적인 기능도 고려합니다.
")
한약재 시스탄체 식물-항종양
2. 암 치료에서 세포사멸의 복잡한 역할: 프로그램화된 세포 사멸
세포사멸 기능장애로 인한 질병의 발병기전을 이해하려면 세포사멸 메커니즘에 대한 이해가 필요합니다. 세포사멸 유전자나 경로를 특별히 표적으로 삼는 약물의 생성은 이로부터 이익을 얻을 수 있습니다. 카스파제는 개시자와 실행자로 기능하기 때문에 세포사멸 메커니즘의 필수적인 부분입니다. 카스파제가 활성화될 수 있는 세 가지 뚜렷한 경로가 있습니다. 내인성(또는 미토콘드리아)과 외인성은 가장 자주 논의되는 두 가지 세포사멸 개시 경로(또는 사멸 수용체)입니다(그림 1). 두 경로 모두 궁극적으로 공통 경로인 세포사멸의 실행 단계로 이어집니다. 내인성 소포체 경로는 잘 알려지지 않은 세 번째 개시 경로입니다[55].

그림 1. 암 치료에서 세포사멸의 역할
세포사멸 실행 단계에서는 수많은 카스파제가 활성화됩니다. 카스파제 9는 내인성 경로를 매개하고, 카스파제 8은 외인성 경로를 매개합니다. 내인성 경로와 외인성 경로 모두 카스파제 3에 모여서 세포사멸을 완료합니다. 핵 세포사멸은 카스파제 3에 의한 카스파제 활성화 데옥시리보뉴클레아제 억제제의 분해로 인해 발생합니다. 추가적으로, 단백질 키나제, 세포골격 단백질, DNA 복구 단백질 및 엔도뉴클레아제 억제 서브유닛은 하류 카스파제에 의해 절단됩니다. 또한 이들은 세포골격, 세포 주기 및 신호 전달 경로에 영향을 미치며, 이들 모두는 세포사멸 중에 발생하는 특정 형태학적 변화를 형성하는 데 도움이 됩니다. 세포사멸은 막 수포 형성, 세포질 염색질 응축 수축 및 핵 붕괴를 특징으로 하는 에너지 의존적 프로그램 세포 사멸입니다. Apoptosis는 염증을 일으키지 않고 세포가 죽는 과정입니다 [57,58]. 또한 미토콘드리아 매개 메커니즘 또는 표면 사멸 수용체(DR; 외인성 세포사멸)(내인성 세포사멸)에 의해 시작될 수 있습니다[59-62]. 두 경로 모두 집행 카스파제가 활성화되어 세포사멸 세포 네트워크의 구조 및 조절 분자와 관련된 분자를 절단합니다[46,63,64]. 병원성 스트레스 요인 이후에 세포사멸은 세포의 항상성을 유지하는 데 도움이 되는 세포 사멸 메커니즘입니다[65]. 악성 세포는 일반적으로 병원성 자극에서 살아남기 위해 일련의 유전적 돌연변이를 겪습니다. 세포사멸 저항성 또는 감소된 세포사멸은 발암을 촉진합니다 [66-68]. 암세포는 세포사멸 촉진 및 항세포사멸 단백질 균형을 재조정하여 세포사멸을 피하는 경우가 많습니다. 암세포는 또한 카스파제 활성이 낮고 DR 신호 전달이 손상된 경우 세포사멸을 피할 수 있습니다[62,69,70]. Bcl-2 계열 단백질, 세포사멸 단백질(IAP) 억제제, p53, 실행자 카스파제 및 DR은 암세포의 영향을 받는 경우가 많습니다. 이러한 분자 유전자와 관련 경로는 세포 사멸 세포 사멸을 유발하기 때문에 암 치료에 중요합니다 [71-75]. 암 치료에서 세포사멸의 전형적인 역할이 묘사되어 있습니다(그림 1).
3. 해초: 황산화 다당류의 주요 원인
엄청난 생물 다양성과 전 세계적으로 식품 및 전통 의학으로 사용되기 때문에 해조류는 생리 활성 화학 물질의 좋은 공급원으로 생각됩니다 [76-78]. 다양한 해조류 유래 생리 활성 화학 물질, 가공되지 않은 추출물 및 부분 정제된 다당류가 다양한 인간 질병에 대한 치료 효과가 조사되었습니다[54,79,80]. 항산화 특성은 해초로 만든 식물산물이 질병에 저항하는 능력을 돕습니다. 갈조류에는 알긴산, 후코이단을 비롯한 다양한 물리적, 기능적으로 구별되는 다당류가 있습니다 [81,82]. 생명공학, 의학 및 식품 제조 분야에서는 황산화 다당류가 자주 사용됩니다[83,84]. 해조류에서 분리된 폴리페놀, 유리 아미노산, 요오드 함유 물질, 비타민, 지질 등은 식품 가공 및 의약에 사용되는 저분자 대사산물의 예입니다[81,85]. 황산화 다당류는 생리학적으로 활성이 있고 고도로 분지형이며 단당류 조성과 다르며 더 높은 분자량을 가지고 있습니다. 연결된 설탕 분자의 긴 사슬이 황산기 [86]로 장식된 후코이단을 구성합니다. 많은 악성 종양을 퇴치하는 황산화 다당류의 능력은 주로 항산화 능력에 기인합니다 [87-89]. 해조류에서 추출한 황산화다당류가 효과적인 항암제라는 사실은 잘 알려져 있습니다. 해초에는 다양한 황산화 다당류가 포함되어 있습니다. 화학적 구성에 따라 다당류는 갈라티아와 황산화 자일란, 황산 다당류(일반적으로 녹조류에서 발견됨)로 분류됩니다. 갈조류에도 후코이단이 포함되어 있습니다 [90]. 홍조류에는 종종 한천, 카라기난, 자일란 및 플로리디안이 포함되어 있습니다. 여러 가지 조류 황산화 다당류가 다양한 인간 건강 불평등을 해결하기 위한 치료 후보로 사용될 수 있습니다[91]. 카라기난으로 알려진 황산화 갈락탄은 식품 및 의약품 산업에서 자주 사용됩니다. 푸칸과 같은 수용성 섬유질은 갈조류에서 발견됩니다. 반면, 붉은 해조류에는 자일란, 플로리디안 전분, 황산화 갈락탄(한천 및 카라기난)과 같은 수용성 섬유질이 풍부합니다[92]. 녹조류에는 또한 자일로스, 갈락토스, 우론산, 아라비노스 및 람노스뿐만 아니라 만난, 자일란, 전분 및 이온성 황산기를 갖는 다당류도 함유되어 있습니다. 다당류에는 다양한 유형의 수용성 및 불용성 섬유가 있습니다[93,94]. 건조 중량에 비해 해조류는 과일과 채소에서 발견되는 것보다 더 많은 식이섬유(25~75%)를 제공합니다[95]. 조류에서 추출한 식이섬유를 섭취하면 항종양, 항암, 항응고제, 항바이러스제 역할을 하기 때문에 건강에 많은 긍정적인 효과가 있습니다. 갈색 대형조류에서는 황산화 다당류가 세포벽에 광범위하게 분포되어 있습니다[80]. 다른 생물학적 작용 중에서 황산화 다당류는 항산화제, 항염증제, 항응고제, 항암제, 항바이러스제, 항당뇨병제 및 항혈전제 역할을 합니다. 그들은 또한 인간의 면역체계를 변화시킵니다[7]. 또한, 갈조류에 풍부하고 황산화 다당류의 두 번째로 큰 공급원인 후코이단은 인체 건강에 있어 장내 대사를 촉진합니다[94].
해조류 유래 황산화 다당류의 구조적 복잡성
연구는 해조류에서 발견되고 항암, 항산화, 항응고 및 항염증 특성을 갖는 다당류에 점점 더 초점을 맞추고 있습니다 [7,96]. 다당류는 단량체 단위에 따라 호모다당류, 호모글리칸, 헤테로다당류 또는 헤테로글리칸으로 분류되는 큰 분자입니다. 다당류는 또한 해초 기원에 따라 갈색, 빨간색, 녹색 또는 파란색으로 분류됩니다. 후코이단(황산화 다당류)은 갈조류의 주성분입니다. 한천, 자일란, 카라기난, 플로리디안 전분(아밀로펙틴과 유사한 글루칸), 수용성 황산화 갈락탄 및 포르피린은 홍조류로 만든 일부 제품입니다. 녹색 해조류에는 황산화 갈락탄, 자일란 및 다당류가 포함되어 있습니다. 해조류에는 다양한 다당류가 포함되어 있으며 Ascophyllum, Porphyra 및 Palmaria를 포함한 일부 속은 건조 중량을 기준으로 최대 76%의 다당류를 함유하고 있습니다[97].
4. 황산화 다당류의 질병 예방 활성: 마법의 총알
수많은 연구를 통해 다당류의 생물학적 활성이 분자량, 형태 상태, 화학적 조성 및 글리코시드 연결에 의해 영향을 받는다는 것이 입증되었습니다[98]. 다당류 점도, 형태, 수용성 등과 같은 필수 특성과 분자량 사이의 관계를 이해하는 것은 암에서 중요합니다[99,100]. 저분자량(LMW)의 포르피린은 더 강력한 항산화 특성을 가지고 있습니다. 포르피란은 평균 분자량이 더 낮기 때문에 ROS 제거 활성이 더 중요합니다[103]. 또한 포르피린산 가수분해의 부산물인 올리고포르피린은 수많은 암을 예방하고 치료할 수 있는 잠재력을 가지고 있습니다. 감마선이 P. yezoensis 유래 포르피린의 항암 반응을 손상시켰기 때문에 더 높은 방사선 노출량과 더 낮은 분자량의 포르피린이 필요했습니다 [45]. 저분자량 포르피린이 더 강력한 항암 활성을 가지고 있다고 주장하는 이전 연구와는 달리, 포르피린은 분해된 생성물보다 암 세포주인 HeLa 및 Hep3B를 더 강력하게 억제했습니다. 단당류나 황산염의 조성은 크게 변하지 않았다[100]. 따라서 향후 연구는 포르피린의 분자량과 항암 활성이 어떻게 관련되는지에 초점을 맞춰야 한다. 일본, 중국, 태국, 한국과 같은 아시아 국가에서는 식용 해초가 건강에 좋은 식품으로 높이 평가됩니다. 암과의 싸움에는 폴리페놀, 테르펜, 피코빌리단백질, 카로티노이드, 플로로탄닌, 색소 및 다당류의 사용이 필요합니다. 해조류의 항암 특성에서 발견되는 항산화제는 암의 확산을 막는 데 도움이 됩니다. 암이 진행됨에 따라 항산화제는 세포 독성을 유발하지 않고 종양 성장을 억제하기 때문에 중요합니다[105]. 예를 들어, 육종 180의 마우스 모델은 세포독성 없이 Champia feldmannii의 면역 자극 황산화 다당류로 성공적으로 치료되었습니다[106]. Gracilaria lemaneiformis의 다당류는 비장세포 증식, 대식세포 식균작용 및 종양 억제를 유도했습니다. H22 간암 세포 이식을 받은 마우스의 혈액 내 IL-2 및 CD8+ T 림프구 수치가 더 높았습니다[107]. C. feldmannii의 황산화 다당류는 스위스 생쥐의 시험관 내 및 생체 내에서 항암 효능을 보여주었습니다. OVA 특이적 항체의 생산을 늘리면 면역력이 향상됩니다[106]. 후코이단의 항암 특성은 위암, 유방암, 폐암, 간암 등 다양한 암에서 확인되었습니다[7]. 후코이단은 포르피린 및 기타 황산화 다당류보다 더 많은 주목을 받아왔습니다. 녹조류, 갈조류, 홍조류의 황산화 다당류는 항암 특성으로 인해 이러한 맥락에서 많은 관심을 불러일으켰습니다. 다양한 황산화 다당류의 물리화학적 특성과 광범위한 치료 잠재력(표 1)이 이 연구를 주도할 것입니다. 이온 용해도, 가교결합, 생체적합성, 무독성, 유변학적 특성 및 생분해성을 포함하여 황산화 다당류의 일부 물리화학적 특성이 보고되었습니다[108-110]. 이러한 특성은 황산화 다당류의 중요한 특성으로 응용 분야에 많은 관심을 불러일으켰습니다. 후코이단의 주요 특성은 이온 가교 및 용해도입니다[109]. 수용성 황산화 다당류는 후코이단 및 기타 양전하를 띤 분자 기반 전달 시스템의 개발을 촉진합니다. 예를 들어, 후코이단의 음전하를 띠는 황산염 그룹은 키토산의 암모늄 그룹과 공유되어 나노입자, 하이드로겔 및 영양식품 전달을 위한 식용 필름을 형성할 수 있습니다[111,112]. 무독성은 이온 가교 및 용해도 외에도 중요한 특성입니다. 이전에 언급한 경우를 제외하고, 생분해성과 생체적합성은 치료 및 약물 전달 시스템에서 황산화 다당류의 사용을 촉진하는 데 중요한 요소입니다. 최근 연구자들은 우수한 생체적합성과 생분해성 때문에 황산화 다당류에 관심을 가지게 되었습니다. 황산화 다당류의 생분해성은 생리 활성 성분의 생체 이용률과 전달 효과를 증가시킬 수 있습니다. 해중합 및 정제는 일반적으로 황산화 다당류의 분자량을 낮춤으로써 생분해성을 증가시킬 수 있으나, 이 방법은 너무 비싸서 널리 사용되지 않는다[113].
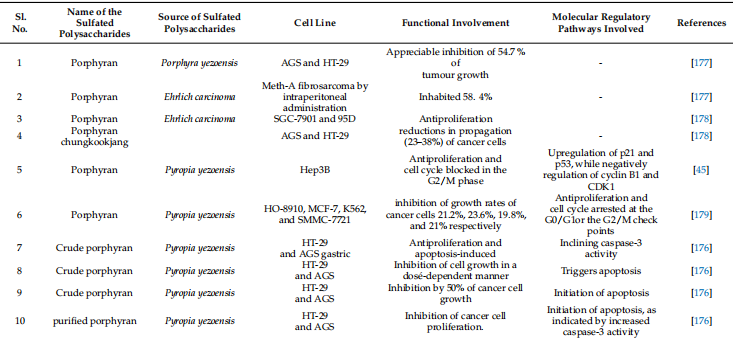
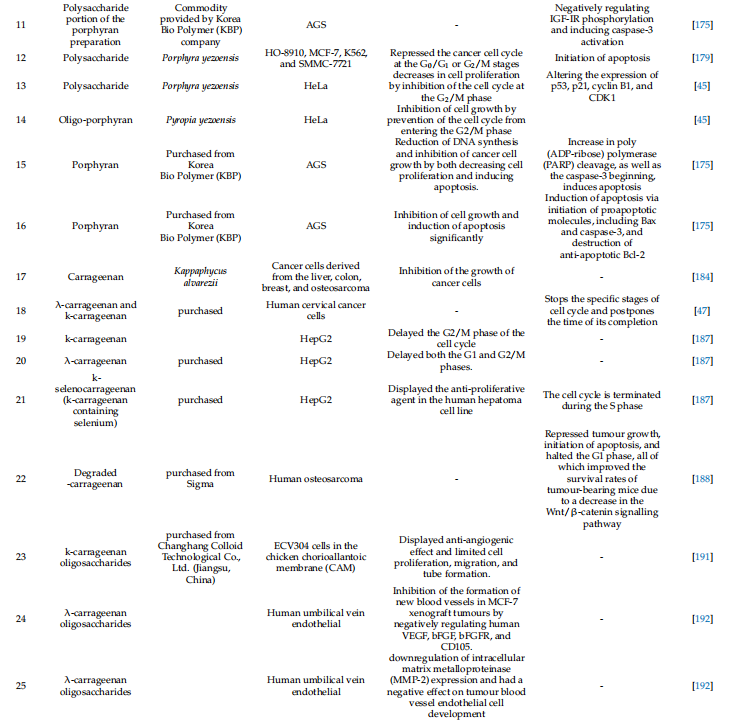
표 1. 황산화 다당류의 물리화학적 특성 및 기타 기능과 함께 치료 가능성

세포사멸 조절 해조류 유래 황산화 다당류
항암 항혈관신생 및 항염증 약물로서 황산화 다당류는 다양한 생물학적 효과를 갖습니다[43,114]. 다양한 해양 서식지에서 분리된 황산화 다당류는 광범위하게 연구되어 수많은 세포 생존 경로를 조절하고 세포사멸을 유도함으로써 다양한 암 세포주에 대한 효과적인 항암 매개체인 것으로 밝혀졌습니다[43,46]. 따라서 다양한 해양 서식지의 다양한 녹색, 갈색, 붉은 해조류에서 분리된 황산화 다당류가 세포 사멸 경로를 동적으로 조절하는 능력에 대해 광범위하게 연구되었습니다. 이러한 다당류는 수많은 세포 생존 경로를 조절하고 세포사멸을 유도하기 때문에 효과적인 항암제입니다. 황산화다당류는 암치료는 물론 정밀의료까지 차세대 의약품 개발을 위해 널리 사용되고 있다. 녹조류: 엽록소라고도 알려진 녹조류는 다당류의 공급원인 영향력 있는 해양 조류 그룹입니다[115]. 그러나 녹조류 세포벽 다당류는 적색(아가란 및 카라기난) 및 갈조류 다당류(후코이단)보다 덜 주목을 받았습니다[96]. 그럼에도 불구하고 녹색 해조류에서 추출한 황산화다당에 대한 연구는 주로 구조적 다양성, 생물학적, 물리화학적 특성으로 인해 최근 과학계의 관심을 불러일으켰습니다[116]. 더욱이, 분지형 구조를 초래하고 다양한 특수 분포를 갖는 황산염 그룹이 부착된 다양한 글리코시드 결합으로 인해 황산화 다당류는 가장 생체 활성이 높고 유망한 후보입니다[117]. 울반(Ulvan)은 수용성이며 녹조류의 세포벽에서 추출되는 황산화 다당류입니다. 그들은 Ulva, Enteromorpha, Monostroma, Caulerpa 및 기타 속에 속하는 식물에 존재합니다. 이는 황산화 람노스와 우론산(글루쿠론산 또는 이두론산)과 같은 반복되는 이당류 부분으로 구성됩니다. 동물 결합 조직의 세포외 기질에 존재하는 글리코사미노글리칸은 Ivan 이당류 부분과 유사한 구조를 가지고 있습니다. 일부는 자일로스 잔기가 눈에 보이기도 합니다(그림 2)[118]. Codium 비용의 고피루베이트 1,3-D-갈락탄 황산염 및 Codium의 이와 유사한 다당류인 thmocladium은 녹조류에서 발견되는 두 가지 다른 유형의 다당류입니다[119,120]. 황산화 -D-만난(Codium vermilara)에서 분리된 것과 같은 것도 발견되었습니다[121]. 반 황산화 다당류의 분자 구조는 그림 2에 표시되어 있습니다.

그림 2. Ivan의 분자 구조는 ChemDraw 12에 그려져 있습니다.0 Ultra.
다양한 열대 녹조류에서 분리된 황산화 다당류는 최근 항산화 및 항증식 특성을 갖는 것으로 밝혀졌습니다. 72시간 배양 후, Caulerpa prolifera에서 분리한 다당류에 의해 HeLa 세포 증식이 36.3%에서 58.4%로 감소했습니다[122]. 녹조류인 Caulerpa racemosa의 두 가지 다당류 분획은 100mg/kg/day의 용량에서 항종양 활성을 나타냈으며, 마우스에 이식된 H22 종양 억제율은 59.5~83.8%(48시간) 및 53.9%(14일)입니다. ), 각각 [123]. 생체 내 및 시험관 내 실험을 통해 Enteromorpha prolifera의 수용성 황산화 다당류 분획이 면역을 자극하는 것으로 밝혀졌습니다. 이러한 다당류는 증가된 m-RNA 발현을 통해 ConA에 의해 유발된 비장세포 증식과 사이토카인 생산을 크게 증가시켰습니다. Ulvariga의 Ulvan은 쥐 대식세포의 분비와 활동을 자극하고 COX{21}} 및 NOS{22}} 발현을 증가시켰으며 일부 사이토카인의 발현을 두 배 이상 증가시켰습니다[125]. Ulva pertusa의 Ulvans는 산화질소와 사이토카인 생성을 자극하는 동시에 종양 세포에 대한 세포독성을 거의 일으키지 않습니다. 쥐의 D-갈락토사민 유발 간염 실험에서 Ivan의 항산화 활성에 대한 여러 연구가 발표되었습니다[127,128]. 녹조류에서 추출한 다당류는 강력한 면역 조절 및 항산화 특성을 갖고 있어 암 예방에 사용될 수 있음을 암시합니다. Ulvan의 항암 활성은 최근 U. australis, U. lactuca, U. ohnoi 및 U. rigita에서 발견되었습니다[129]. 여러 연구에서 ulvan의 항암 활성, 특히 항유방암, 항결장암 및 항자궁암 활성을 테스트하기 위해 독성 및 세포 생존력에 대해 조사했습니다[129-132]. Ulvan에는 간세포 암종의 증식을 억제하고 세포 사멸을 유도하는 황산화 다당류가 포함되어 있습니다. 산화 스트레스를 낮춤으로써 황산화 다당류는 DNEA로 인한 손상으로부터 간을 보호합니다[133]. 또한 DMBA로 치료한 생쥐에서 세포사멸을 강화하고 산화 스트레스와 염증을 감소시키며 항산화 방어 시스템을 강화합니다. Ulvan은 A459 및 LS174 세포에 대한 독성이 낮았지만(IC50 > 200mg/mL), Fem-x 및 K562 세포(각각 IC50 74.73 및 82.24mg/mL)에 대해서는 더 효과적이었습니다. 중간 정도의 세포 독성을 예방합니다 [134]. 21~99μg/mL 범위의 IC50 값을 사용하면 MCF-7 및 HCT-116 세포에서 종양 성장이 감소하고[132] 강한 리간드 결합이 이를 황산화 다당류에 연결하는 것으로 보입니다[135]. Ulvan은 간세포암종(IC50 29.67 ± 2.87 µg/mL), 인간 유방암(IC50 25.09 ± 1.36 µg/mL) 및 자궁경부암(IC{{65)의 성장을 억제했습니다. }}.33 ± 3.84 µg/mL) [131]. 그러나 Caco{70}} 세포 증식 또는 분화는 저분자량 다당류(5000 Da), 일반적으로 올리고당에 의해 억제될 수 있습니다[129]. 황산화 다당류는 항증식 효과가 있지만 세포 유형에 따라 다릅니다. 녹조류 Codium Bernabei에서 추출한 황산화 다당류 TP(알코올에 침전)는 AP(산성 배지에 침전)와 비교하여 HCT-116 및 MCF-7 세포주에서 낮은 세포독성을 나타냈습니다. 반면, HL-60 세포주는 AP에 노출되었을 때 세포독성이 거의 나타나지 않았습니다[51]. 강력한 항산화 활성으로 인해 Enteromorpha spp. 추출물은 Fem-x, A549, LS174 및 K562와 같은 암 세포주에 항증식 효과가 있습니다. 또한 Enteromorpha 압축 추출물의 다른 용매 추출물은 구강암 세포주 Cal33 및 FaDu에서 세포사멸을 통해 항암 활성을 유도합니다. 갈조류: 갈조류는 황산화 다당류의 가장 유망한 공급원이며 다양한 암 세포주에 대해 가장 유망한 항암 활성을 나타냅니다. 갈조류에서 유래된 전형적인 황산화 다당류 구조가 표시됩니다(그림 3). 루이스 폐암 세포(LCC)와 흑색종 B16 세포는 Sargassum sp.에서 분리된 후코이단에 민감한 것으로 밝혀졌습니다. [137]. 이는 형태학적 변화에서 볼 수 있듯이 세포 증식을 감소시키고 용량 의존적으로 세포사멸을 촉진했습니다. S. hemiphyllum의 후코이단은 miR-29c를 상향 조절하고 miR-17-5p를 하향 조절하여 유방암의 성장을 억제했습니다. 또한, 후코이단 투여 후 E-카드헤린의 증폭과 N-카드헤린 발현 감소로 인해 EMT 진행이 느려지는 것이 분명했습니다. 더욱이, phosphoinositide 3- 키나아제/Akt 경로의 활성화는 유방암 세포에서 세포사멸을 촉진했습니다[138]. L. gurjanovae의 후코이단은 EGFR 인산화를 지연시켜 쥐 표피 JB6 Cl41 세포에서 항종양 효과를 입증했습니다. 이는 EGF에 의해 유발된 c-jun 신호 전달을 제어하고 활성화 단백질-1(AP{100}}) [139]의 작용을 억제했습니다.

그림 3. 잠재적인 치료 효과가 있는 후코이단과 같은 다양한 유형의 황산화 다당류의 분자 구조가 ChemDraw 12에 그려져 있습니다.0 Ultra.
F. vesiculosus에서 추출한 후코이단은 난소암(ES2 및 OV90) 세포에서 세포 증식을 억제하고 세포 주기를 정지시켰습니다. 또한 내인성 세포사멸을 조절하는 ROS도 생성했습니다. PI3K 및 MAPK 신호 전달 경로를 억제함으로써 ER 스트레스는 또한 세포 사멸을 촉진했습니다. 또한 p-38 MAPK, ERK1/2 및 JNK 경로(MC3)를 변형하여 인간 점액표피암종에 대한 항암 효과를 입증했습니다[140]. 또한 세포질과 미토콘드리아의 칼슘 양을 줄여 세포 사멸 세포 사멸을 지원했습니다. F. vesiculosus에서 생산된 후코이단에 대한 유사한 추출 기술은 제브라피시 이종이식 및 fli1 Tg 모델에서 생체 내 항암 효능을 보여주었습니다[141]. HepG2 및 HeLa G-63 세포에서 Fucus vesiculosus의 후코이단은 강력한 항암 활성을 나타냈습니다. 후코이단은 인간 간암 세포(HepG2)에 더 효과적인 것으로 밝혀졌습니다[142]. Fucus vesiculosus의 후코이단은 인간 버킷 림프종(HS-Sultan) 세포에서 카스파제{14}}의존성 세포사멸을 유도하는 MMP를 증가시켰습니다. 또한, 후코이단 투여 후 HS-술탄 세포에서 카스파제 독립적 세포사멸 세포사멸이 보고되었습니다. 또한 후코이단은 세포사멸 활성화에 필요한 ERK 및 GSK 경로의 인산화를 방지했습니다. 낮은 IC50(34 µg/mL)은 HCT에서 Bcl-2을 하향 조절하면서 프로-카스파제-3, 프로-카스파제-9 및 카스파제-3/7을 활성화했습니다.{{28 }} 셀 [144]. Bcl-xl, Bcl{32}} 및 Mcl{33}}과 같은 항세포사멸 단백질이 MDA-MB231 세포에서 세포사멸을 일으키는 능력은 IC50(820μg/mL)의 후코이단에 의해 억제되었습니다[46]. 후코이단(IC50; 20 µg/mL) 치료는 결장암 세포주에서 Bad, Bcl{41}}, Bim, Bcl-xl 및 Bik의 발현에 유사한 변동을 가져왔습니다[144]. C. okamuranus에서 유래한 후코이단은 Con A와 결합하여 HL60 세포에서 카스파제{44}}/7 유도에 의해 내인성 세포사멸을 촉진했습니다[145]. 또한, 글루타티온 고갈과 NO 생성은 MEKK1, ERK1/2, MEK1 및 JNK의 활성화와 마찬가지로 인간 백혈병 세포에서 세포사멸의 중요한 중재자였습니다. C. novaecaledoniae의 후코이단을 추출하여 HeLa, MCF-7, MDA-MB-231 및 HT1080 세포에서 내인성 세포사멸을 유도하는 데 사용했습니다. 이러한 내인성 세포사멸은 MMP, DNA 단편화, 핵 응축 및 포스파티딜세린 외부화를 동반했습니다. C. 오카무라누스 후코이단은 카스파제-3 및 -7 경로를 유도하여 U937 세포에서 카스파제 의존성 세포사멸을 유도했습니다[148]. 또한 생체 내 모델에서 마우스 세포 매개 면역, 식세포 및 면역 세포 증식을 강화했습니다 [149].
")
cistanche 보충 혜택 - 면역력 증가
또한 정상 위(Hs 677. St) 세포에서 C. okamuranus에서 분리한 후코이단은 5-플루오로우라실(5- FU)[150]로 인한 세포 손상을 감소시켰습니다. 이 설정에서는 인간 유방 상피 세포에 대한 세포독성 없이 MCF-7 세포에서 상당한 항증식 활성이 관찰되었습니다. 두 세포주 모두에서 카스파제-7, 카스파제-8 및 카스파제-9 활성, 뉴클레오솜간 DNA 단편화 및 염색질 응축이 증가했습니다[151]. 후코이단 요법은 MCF-7 [152]에서 카스파제 독립적인 세포 사멸에 대해서도 보고되었습니다. 가수분해는 관강 후코이단 함량을 증가시키며, 이는 이러한 후코이단을 소화하지 않기 때문에 결장암의 강력한 화학적 예방 매개체입니다[153]. 후코이단(0~20 µg/mL) 치료법은 카스파제-3 조절을 통해 HT{16}} 및 HCT116 세포에서 미토콘드리아 사멸을 촉진했습니다. HT-29 세포의 외인성 세포사멸도 최근 보고되었습니다[144]. C. 오카무라누스 저분자량 후코이단(LMWF; 6.5~40kDa), 고분자량 후코이단(HMWF; 300~330kDa), 중간분자량 후코이단(IMWF; 110~138kDa)의 항암 특성이 한 연구에서 입증되었습니다. 결장암 종양 보유 쥐 모델 [154]. U. pinnatifida의 후코이단(MW 5100 kDa)은 ERK1/2 MAPK 유도, p38 MAPK 억제 및 PI3K/Akt 경로를 통해 인간 전립선암(PC{31}}) 세포에서 세포사멸을 유도했습니다. 더욱이, Wnt/-catenin 경로의 하향조절은 세포사멸의 진행을 도왔습니다. 후코이단은 또한 PC{40}} 세포의 p21Cip1/Waf 경로를 증폭시켰습니다. 또한 E2F-1 세포주기 관련 단백질을 감소시키는 동시에 Wnt/-catenin 경로를 증가시켰습니다. GSK-3 활성화는 c-MYC와 cyclin D1의 발현을 감소시켜 항증식 활성을 도왔습니다[156]. 이러한 후코이단은 이전에 언급한 중요한 세포 신호 전달 경로를 변경하여 HeLa, A549 및 HepG2 세포에서 항암 활성을 갖는 것으로 밝혀졌습니다[157]. F. vesiculosus의 후코이단은 NB4, THP-1 및 HL-60을 포함한 암 세포주에서 세포사멸을 유도했습니다. 후코이단 투여는 caspases-3, -8 및 -9를 활성화하고 Bid를 절단하고 HL-60 세포에서 MMP를 변경했습니다. 세포사멸의 시작은 U937 세포에서 비슷한 효과를 나타냈습니다. 또한 U937 세포에서 후코이단 치료는 Bax/Bcl-2 비율뿐만 아니라 MMP(미토콘드리아 막 전위)와 세포질 시토크롬 C 방출을 증가시켰습니다. 반면, 카스파제 억제제는 세포사멸의 시작을 지연시켜 후코이단 조절 카스파제 활성이 세포사멸 유도에 영향을 미친다는 것을 입증했습니다. 더욱이, 특정 p38 MAPK 억제제인 SB203580을 사용한 치료는 세포사멸 할인에 대한 원인이 되었으며, 이는 세포사멸 활성화에서 MAPK의 중요성을 입증했습니다. 후코이단 요법은 사이클린 D1, 사이클린 E 및 Cdks(사이클린 의존성 키나제)에 영향을 주어 EJ 세포의 G1 세포 주기를 억제했습니다. 또한 Rb 인산화를 억제하여 세포 노화를 유발합니다. F. vesiculosus의 후코이단은 G1 단계에서 세포주기를 멈추고 CDK-4 및 cyclin D1 수준을 낮추어 MCF-7 세포의 성장을 억제했습니다. 또한 PARP와 Bid를 절단하고 Bcl-2를 감소시키고 Bax를 증가시킴으로써 ROS 의존성 세포사멸을 유도했습니다. MCF-7 세포는 caspase-7, -8, -9 및 세포질 시토크롬 C 방출의 조절을 통해 내인성 세포사멸의 시작을 나타냈습니다[160,161]. 또한, F. vesiculosus 치료의 후코이단은 MMP-9를 하향 조절하고 E-cadherin을 과발현함으로써 MCF-7 세포에서 EMT뿐만 아니라 세포 이동과 침입을 감소시켰습니다. F. vesiculosus에서 추출한 후코이단은 MDA-MB-231 및 4T1 이종 이식 암컷 Balb/c 마우스 세포의 성장을 억제하여 전이성 폐 결절 발달을 줄였습니다. TGFR 유도 EMT의 효과적인 좌절은 TGFRII 및 TGFRI를 하향 조절함으로써 기계적으로 달성되었습니다. 위에 언급된 사례는 모두 상피 마커의 상향 조절 및 Smad2/3 Smad4 발현의 인산화, Smad2/3 Smad4 발현의 인산화 및 하류 신호 전달 분자와 관련이 있습니다[163]. 또한, 카스파제-3 활성화, 세포질 시토크롬 C 방출, Bcl-2의 하향조절 및 Bax 발현 증가가 세포사멸을 유도했습니다. 또한, VEGF, Survivin 및 ERKs 발현의 조절은 세포사멸의 시작을 도왔습니다. MDS/AML 및 SKM1 세포주에서 시판되는 후코이단(48시간 동안 100μg/mL)으로 처리하면 세포 주기 정지(G1 단계)와 Fas 자극이 카스파제 8 및 9 변조를 통해 외인성 세포사멸을 유도했습니다. 또한 이는 ROS 의존적 방식으로 PI3K/Akt 경로에 영향을 미쳐 세포사멸을 촉진했습니다. 이는 DU{117}} 세포(전립선암)에서 MAPK 및 PI3K/Akt 신호 전달 경로를 조절하기 위해 p-Akt, p-PI3K, p-P38 및 p-ERK를 변경했습니다. 또한 Bcl-2, PARP 절단 및 caspase-9 발현은 농도 의존적으로 감소하는 반면 Bax 발현은 증가했습니다[166]. 후코이단 투여는 골육종(MG{122}}) 세포에서 세포사멸을 유도했습니다(세포 폭발, 핵 붕괴 및 염색질 응축으로 입증됨). 시판되는 합성 후코이단을 사용한 치료는 미토콘드리아 막 전위(MMP)를 활성화하고 Bax/Bcl{126}} 비율을 증가시키며 세포질 시토크롬 C 방출을 증가시킴으로써 인간 방광암(5637) 세포에서 ROS 조절 세포사멸을 증가시켰습니다. 더욱이, PI3K/Akt 신호전달 및 항텔로머라제 활성의 억제는 텔로머라제 활성을 하향조절함으로써 5637 인간 방광암 세포에서 세포사멸 세포 사멸을 촉진했습니다[168]. 더욱이, AKT 신호전달 활성화는 증식을 억제하고 방광암 세포의 이동 및 침입 능력을 억제하는 데 중요한 역할을 한다고 주장되었습니다[169]. 후코이단은 p21/WAF1, 사이클린 및 CDK의 발현을 변경하여 5637 및 T{133}} 세포(인간 방광암종)의 세포주기를 억제했습니다. 더욱이, AP-1 및 NF-kB를 통한 MMP-9 억제는 방광암 세포 증식을 감소시켰습니다[169]. 강력한 항암제인 갈조류의 황산화 다당류가 표 2에 나와 있습니다.
표 2. 강력한 항암제인 갈조류의 황산화 다당류. ↑: 상향 조절, ↓: 하향 조절, ⊥: 억제



붉은 해초: 포르피란(Porphyran)은 붉은 해초인 Porphyra sp.에서 발견되는 중합체입니다. 포르피란은 L-갈락토스 6-O-황화 및 6-O-메틸화로 크게 대체된 갈락토스입니다[100]. 포르피란의 전형적인 반복 구조가 표시됩니다(그림 4). 포르피란은 홍조류에서 뜨거운 물 추출, 초음파 처리 및 라디칼 분해를 통해 추출됩니다. 인간 연구에서는 포르피란의 항암, 고지혈증 및 항염증 특성이 입증되었습니다[170]. 포르피란을 경구로 섭취하면 ICR 생쥐의 간을 고지방 식이의 영향으로부터 보호하여 식이 저지혈 성분으로 사용될 수 있음을 시사합니다[171].

그림 4. 잠재적인 치료 효과가 있는 다양한 유형의 포르피란의 분자 구조가 Chemdraw 12에 그려져 있습니다.0 Ultra.

그림 5. 잠재적인 치료 효과가 있는 다양한 유형의 카라기난의 분자 구조가 ChemDraw 12에 그려져 있습니다.0 Ultra. Υ-카라기난과 같은 카라기난의 다양한 분자 구조의 고분자 구조(a); -카라기난(b); δ-카라기난(c); -카라기난(d); μ-카라기난(e); κ-카라기난(f); ν-카라기난(g); ι-카라기난(h); λ-카라기난(i); 및 θ-카라기난(j).
암은 활성산소와 ROS(활성산소종)에 의해 가속화되는 것으로 알려져 있습니다. 합성 화학예방 약물은 일반적으로 낮은 선택성과 광범위한 생체분포로 인해 종양 환경에서 바람직하지 않은 부작용을 발생시킵니다. 포르피린은 세포 증식, 세포 주기 및 세포사멸 유도에 영향을 미치기 때문에 강력한 화학예방제입니다. 홍조류 Porphyra yezoensis는 시험관 내에서 암세포주에서 세포사멸을 유도할 수 있지만 정상 세포에는 세포독성을 일으키지 않습니다. 일반적으로 포르피린은 건강한 세포에는 독성이 없지만, 암세포에는 독성이 있어 용량 의존적으로 세포 사멸을 초래합니다[175]. 또한, 포르피린은 AGS 인간 위암 세포에서 세포사멸을 유도하면서 전반적인 세포 성장을 억제한다는 것이 입증되었습니다[175]. AGS 세포에서 인슐린 유사 성장 인자-I 수용체/Akt 경로는 PARP 절단 및 카스파제{7}} 활성화를 증가시켜 세포 사멸을 촉진합니다. 수많은 연구에서 포르피린과 그 올리고당의 항종양 및 항암 특성이 입증되었습니다. 포르피린은 위암 세포에서 폴리(ADP-리보스) 중합효소의 절단과 카스파제 3의 활성화를 촉진할 수 있습니다. 포르피린은 AGS 세포(위암)의 발현 수준을 감소시킴으로써 암세포의 성장을 늦출 수 있습니다. 그러면 IGF-IR 인산화가 방지되고 카스파제 3이 활성화됩니다. 조질 및 정제 포르피린은 시험관 내 HT{14}} 및 AGS 세포에서 항증식 활성을 나타냅니다. 세포사멸은 카스파제-3 활성화의 증가로 나타난 바와 같이 조 포르피린 다당류 성분에 의해 유도됩니다[176]. 포르피린은 카스파제-3 [176]를 활성화하여 HT{17}} 세포 증식을 억제합니다. 포르피린은 마우스 종양 모델에서 Ehrlich 세포(EAC) 암종 및 Meth-A 섬유육종에 대해 효과적인 것으로 나타났습니다[177].

사막 생활 시스탄체세뇨관
【추가 요청】 이메일:cindy.xue@wecistanche.com / Whats App: 0086 18599088692 / Wechat: 18599088692
천연 포르피린은 MDA-MB-231에 영향을 미치지 않는 것으로 밝혀진 반면, 감마선 조사에 의해 생성된 포르피린과 두 개의 OP(올리고-포르피린)를 항암 활성에 대해 테스트할 때 두 가지 분해 생성물이 영향을 미쳤습니다. 세포 주기가 G2/M 단계로 진입하는 것을 방지함으로써 OP는 세포 성장을 감소시키는 능력을 갖습니다[45]. 결과적으로 포르피란의 MW는 항암 효능에 상당한 영향을 미치는 것으로 나타났습니다. 낮은 MW의 OP는 암에 특히 효과적입니다. 그러나 거대분자 포르피린에는 항종양 활성이 없습니다. 또한 포르피린의 항암 활성이 밝혀졌는데, 포르피린은 주로 세포 성장을 방해하고 세포사멸을 유도하여 항암제로 작용합니다[180]. 카라기난은 시험관 내에서 암 세포주에서 항증식 활성을 갖고 생쥐에서 종양 성장 억제 효과를 갖는 것으로 수많은 연구에서 나타났습니다[181-183]. 또한 암세포가 기저막과 연결되는 것을 방지하고 종양 세포의 증식과 다른 기질에 대한 접착을 제한함으로써 항전이 효과도 있습니다. 그러나 정확한 작용 메커니즘은 아직 알려져 있지 않습니다. Kappaphycus alvarezii의 카라기난은 간, 결장, 유방 및 골육종에서 암세포의 성장을 예방하는 것으로 밝혀졌습니다. Yamamotoet al. (1986)은 다양한 해조류를 경구 복용하면 생체 내 발암 발생이 크게 감소한다는 사실을 발견했습니다[185]. Hagiwaraet al. (2001) [186]은 수컷 쥐의 결장 발암에 대한 카라기난의 영향을 조사했습니다. 치료는 임상 증상이나 체중에 영향을 미치지 않았습니다. 조직학적 연구에 따르면, 카라기난은 기존 실험 환경에서 최대 식이 섭취량 5.0%에서 대장암 유발 활성이 없습니다[186]. 카라기난은 암세포에 특정 세포독성 효과를 갖는 것으로 여러 연구에서 나타났습니다. 그러한 연구에서 k-카라기난과 람다-카라기난 둘 다 250-2500μg/mL의 용량은 특정 단계에서 세포 주기를 중단하고 완료를 지연시켜 인간 자궁경부암 세포를 억제했습니다[47]. k-카라기난은 세포 주기(G2/M 단계)를 지연시킨 반면, λ-카라기난은 G1 및 G2/M 단계를 모두 지연시켰습니다. 그러나 k-셀레노카라기난(셀레늄을 함유한 셀레노카라기난)은 인간 간암 세포에서 세포 증식을 억제합니다. 세포주기는 세포주기의 S 단계에서 종료됩니다 [187]. 그러나 생체 내 및 시험관 내 연구에서는 천연 카라기난이 인간 골육종 세포주에서 뚜렷한 항증식 효과를 나타내지 않는 것으로 나타났습니다. Wnt/-카테닌 신호 전달 경로의 감소로 인해 분해된 카라기난에 의해 유도된 세포사멸은 종양 성장을 억제하고 세포 주기의 G1 단계를 중단시켰으며, 이 모든 것이 종양 보유 마우스의 기존 비율을 증가시켰습니다. 혈관신생은 암 진행의 중요한 단계입니다. 결과적으로, 암 치료에서 항혈관신생 활성이 광범위하게 연구되고 있습니다. 카라기난은 수라민보다 항혈관신생 활성이 더 높기 때문에 혈관신생 억제제입니다[189,190]. CAM 모델(닭 융모요막)에서 ECV304 세포에 대한 k-카라기난 올리고당의 항혈관신생 결과는 세포 증식, 이동 및 관 형성을 제한하는 것으로 입증되었습니다[191]. 또한, 올리고당은 인간 bFGFR, bFGF, CD105 및 VEGF를 부정적으로 조절함으로써 MCF{44}} 이종이식 종양에서 새로운 혈관의 형성을 억제했습니다. 인간 제대 정맥 내피 세포는 상대적으로 낮은 농도(150~300μg/mL)의 람다-카라기난 올리고당으로 처리되었으며, 이는 종양 혈관 내피 세포의 발달에 부정적인 영향을 미쳤습니다. 황산화의 양과 위치, 분자량은 황산화 다당류의 생물학적 활성에 영향을 미칩니다. 즉, 화학적 변화는 탄수화물의 생물학적 활성을 변화시킵니다[193]. 예를 들어, 람다-카라기난은 다양한 분자량을 갖는 5가지 다른 화합물로 분해될 수 있으며, 이들 모두는 면역 조절로 인해 항암 특성을 갖고 있습니다. 분자량이 15 및 9.3 kDa인 제품과 같은 저분자량 제품은 탁월한 항암 및 면역 조절 특성을 입증했습니다. 황산화, 아세틸화 및 인산화는 Kappaphycus striatum의 k-카라기난 올리고당의 항암 및 면역 조절 특성을 향상시켰습니다. 화학적 변형은 k-카라기난 올리고당의 산화 활성도 증가시켰습니다. 홍조류의 황산화 다당류와 암 치료제에서의 세포사멸 조절은 표 3에 나와 있습니다. 세포사멸 유도는 화학적 예방제에 의해 채택되는 메커니즘입니다. 다양한 해초에서 유래된 다양한 황산화 다당류는 다양한 암세포주에서 세포사멸을 유발합니다(그림 6). 황산화 다당류는 암에서 다양한 화학적 예방 역할을 나타냈습니다(그림 7).

그림 6. 암 예방에 있어서 다양한 해초에서 유래된 다양한 황산화 다당류에 의한 세포사멸 조절.

그림 7. 황산화 다당류는 암에서 다양한 화학적 예방 역할을 나타냅니다.
표 3. 암 치료에서 홍조류의 황산화 다당류와 이들의 세포사멸 조절
5. 황산화 다당류를 이용한 나노입자 합성과 암 치료 효능에 미치는 영향
현재 이용 가능한 세 가지 주요 암 치료법은 수술, 화학 요법, 방사선 요법입니다. 그러나 화학요법은 건강한 정상 세포에 해를 끼칠 수 있는 수준으로 인해 최근 몇 년간 암 치료의 주류가 되지 못했습니다. 나노입자는 암세포만을 다루기 위한 대체 기술로 등장하여 건강한 세포를 위험으로부터 보호하면서 암세포에 대한 약물의 획득 가능성을 높였습니다[195]. 해조류는 천연 황산화 다당류의 일반적인 공급원이지만 다른 공급원도 있습니다. 상처 관리, 조직 공학, 약물 전달 및 바이오 센서에서 Ivan, 카라기난, 포르피린, 후코이단 및 기타 파생물에 대한 수많은 생물학적 및 생의학적 응용이 조사되었습니다[196]. 해초 다당류는 카르복실기, 수산기, 황산염과 같은 친수성 표면 그룹을 갖고 있기 때문에 생물학적 조직과 쉽게 상호작용합니다. 효율적인 약물 전달 시스템을 위해 원하는 특성을 가진 황산화 다당류 나노입자를 생산하는 준비 기술이 많은 주목을 받았습니다[198,199]. 이온 겔화는 일반적으로 황산화 다당류 나노입자를 생성하는 간단하고 부드러운 공정입니다. 그러나 원하는 모양의 울반, 후코이단, 포르피린, 카라기난 기반 나노입자를 생성하려면 공정 최적화가 중요합니다. 최적화는 pH, 온도, 칼슘이온 농도, 황산화다당류 농도, 첨가속도, 교반속도 등을 조절하여 수행할 수 있다. MCF7과 HepG2 세포 모두 카스파제-8 및 카스파제-9 수준의 증가로 인해 나노입자 알부민에서 반(van)에 의한 증식이 억제되는데, 이는 세포사멸의 유도를 나타냅니다[129]. 항암 치료를 위한 약물 전달 시스템으로 사용되는 금나노입자(AuNps)를 만들 때 포르피린을 환원제로 사용할 수도 있습니다. 예를 들어, 인간 신경교종 세포주는 포르피린(LN{11}})으로 코팅된 AuNps에 더 독성이 있습니다. 그 결과, 포르피린으로 덮인 AuNps가 개발되어 독소루비신 염산염 항암 약물 전달체로 사용되었습니다[200]. 5-플루오로우라실(5-FU)이라고 불리는 티미딜레이트 합성효소 억제제는 오랫동안 암 치료에 사용되어 왔지만 부작용 때문에 사용이 제한되었습니다[201]. 전구약물 5-FU에 대한 수용성 거대분자를 생성하기 위해 포르피린으로 덮인 AuNps를 약물 운반체로 사용하여 5-FU 방출을 지연시키고 부작용을 최소화할 수 있습니다[202]. 포르피란으로 덮인 AuNP는 시험관 내 세포독성 연구에서 안전한 것으로 밝혀졌으며, 이는 약물 전달 시스템으로 사용될 수 있음을 시사합니다[203]. 이 때문에 포르피린을 약물 전달을 위한 환원제 운반체로 사용하면 불리한 효과가 없으며 항암 약물의 효과가 더 빨리 나타날 수 있습니다. 특히 후코이단 포르피란과 카라기난 기반 나노입자는 항암제 전달에 대해 철저히 조사되었습니다(표 4).
표 4. 황산화 다당류를 이용한 나노입자 합성과 암 치료 효능에 미치는 영향

6. 황산화 다당류 연구의 한계와 암 예방 분야의 향후 확장
황산화 다당류는 다양한 의약 용도로 사용되지만 생체 이용률이 낮아 일상 생활에서 사용하기가 비현실적입니다. 서로 다른 황산화 다당류 구조는 서로 다른 기관에 얼마나 잘 흡수되는지에 영향을 미칩니다[7]. 또한 시험관 내 및 생체 내 적용 모두에서 유효 용량의 지속적인 변동으로 인해 임상 시험이 손상됩니다[7]. 황산화 다당류의 시험관 내 효과는 전임상 또는 임상 연구에서 종종 재현되지 않습니다[215]. 또한 세포 내 대사가 느리고 용해도가 제한되어 임상 적용이 더욱 어려워집니다 [216]. 더욱 중요한 것은 이들의 광범위한 치료 적용이 세포 특이성과 분자 표적 선택성의 결과라는 것입니다. 세포, 조직 및 종양 설정에 따라 이러한 생리활성 화학물질은 세포 사멸을 유발하는 방식이 다릅니다. 또한, 임상 연구는 단일 특이적 및 다중 특이적 작용 기능을 이해할 때 더 성공적입니다[216]. 황산화 다당류의 합성 유사체는 생성되고 테스트되면 생물학적 이용 가능성이 더 높아질 수 있습니다[217]. 생체 이용률과 표적 특이성을 높이기 위해 황산화 다당류와 그 합성 등가물은 마이크로 에멀젼, 나노 캐리어, 폴리머, 리포솜 및 미셀을 사용하면 이점을 얻을 수 있습니다[218]. 우리 의견으로는 이러한 기술은 다당류 기반 나노입자를 생성하는 데 앞으로 더 자주 사용될 것입니다. 생체 이용률이 증가된 항암제 전달 측면에서 해조류 다당류 기반 나노입자는 유망한 결과를 입증했습니다[170]. 이러한 기술은 또한 숙주 시스템의 대사와 용해도를 향상시킵니다[170]. 또한 세포사멸의 전임상 및 임상 효능은 표적 특이성에 의해 향상될 것입니다. FDA 승인을 받은 약물과 황산화 다당류를 결합하면 임상 효과가 크게 증가할 수 있습니다[170]. 또한, 황산화 다당류는 식품에 첨가되거나 보조제로 사용될 때 현대 약물의 치료 효능을 향상시킵니다[170].

cistanche tubeulosa-항종양의 장점
7. 결론 및 향후 전망
현재의 암 치료 시스템은 황산화 다당류를 다양한 악성 종양에서 다양한 화학요법 효과가 있는 생체 활성 약물 분자를 발견하기 위한 신뢰할 수 있는 공급원으로 확인했습니다. 최근 몇 년간 FDA 승인 약물 중 절반 이상이 해양 자원에서 직접 추출되었거나 화학적 대응물을 사용하여 생성되었습니다. 해양 자원에서 이러한 황산화 화합물을 분리하고 사용하면 생체 이용률이 더 높고 화학적 구성이 다양하며 비환원성 세포 독성이 있습니다. 이러한 특성으로 인해 해조류 유래 황산화 다당류는 다양한 악성 종양을 치료하는 데 있어 납 약물단으로 작용할 수 있습니다. 그러나 이들의 약학적 활용에 대한 중요한 장벽은 생체 이용률, 향상된 분리, 분리물의 청결성, 단일 약물 다중 표적 특이성 및 세포/조직/암 상황으로서의 표적 선택성입니다. 또한, 이들은 다양한 치료 중재, 저렴한 비용의 상업적 생산, 유망한 전임상 및 임상 적용으로 인해 약물 매개체로서 중요한 역할을 합니다. 한편, 유기체의 광범위한 현장 및 외부 수확과 저렴한 재배 유지 비용으로 인해 해양 해조류에서 이러한 황산화 다당류를 상업화하는 것에 대한 낙관론이 있습니다. 추가적으로, 화학요법을 위한 이들 황산화 다당류의 대규모 제조는 이들 다당류의 화학적 합성의 범위 외 적용에 의해 더욱 효과적이게 됩니다. 다양한 암에 대한 혁신적인 약리학적 제제로서 해조류로부터 황산화 다당류를 분리하고 스크리닝하기 위한 새로운 전망이 출현함에 따라, 그러한 예상 제제의 화학요법적 사용이 곧 번성할 것으로 보입니다. 또한, 나노입자 매개 황산화다당체 기반 나노입자는 지속적인 약물 방출, 높은 안정성, 생체적합성 등이 가능해 향후 임상시험에 활용될 예정이다. 표적화 부분은 바람직하지 않은 부작용을 최소화하면서 다당류 기반 나노입자의 치료 효능을 증가시킵니다. 또한, 이러한 약물 후보를 생성하면 현재 사용 가능한 약물을 개선하여 맞춤형 및 정밀 의학의 발전을 이룰 수 있습니다.
참고자료
1. Younossi, ZM; 코리, KE; 비알코올성 지방간 질환 관리에서 체중 감량을 달성하기 위해 식이요법과 운동을 사용한 생활습관 수정에 대한 임JK AGA 임상 실습 업데이트: 전문가 리뷰. 위장병학 2021, 160, 912-918. [상호참조] [PubMed]
2. Blix, H. 핵 비확산 검증: 미래 확보. IAEA 황소. 1992, 34, 2–5.
3. 세계보건기구. 2018년 알코올 및 건강에 관한 글로벌 현황 보고서; 세계보건기구: 스위스 제네바, 2019.
4. 에드워즈, BK; 아무도 없어요, 오전; 마리오토, AB; Simard, EP; 보스코, FP; 헨리, SJ; 제말, A.; 조현수; 앤더슨, RN; Kohler, BA 연례 보고서(1975~2010년), 폐암, 대장암, 유방암 또는 전립선암 환자의 동반질환 유병률과 생존에 미치는 영향을 다루고 있습니다. 암 2014, 120, 1290-1314. [상호참조] [PubMed]
5. Delgermaa, V.; 다카하시, K.; 박은경; 르, GV; 하라, T.; Sorahan, T. 1994년과 2008년 사이에 세계보건기구에 보고된 글로벌 중피종 사망. Bull. 세계보건기구. 2011, 89, 716-724. [상호참조] [PubMed]
6. 프라단, B.; 나약, R.; 파트라, S.; 지트, BP; Ragusa, A. 산화 스트레스 관련 인간 질병에 대한 강력한 약물 전달 물질로서 해조류의 생체 활성 대사 물질: 종합적인 검토. Molecules 2020, 26, 37. [CrossRef]
7. 프라단, B.; 파트라, S.; 나약, R.; 베헤라, C.; 대시, SR; 나약, S.; 사후, BB; 부티아, SK; Jena, M. 인간의 건강과 질병에 대한 후코이단, 황산화 다당류의 다기능적 역할: 강력한 치료제를 추구하는 바다 밑 여행. 국제 J. Biol. Macromol. 2020, 164, 4263-4278. [상호참조]
8. 파트라, S.; 볼, CS; 파니그라히, DP; 프라하라지, PP; 프라단, B.; 예나, M.; Bhutia, SK 감마 조사는 NRF2 불활성화 신호 전달 경로에서 세포사멸을 유발하는 자가포식 플럭스의 감쇠를 통해 갈산의 화학 감작 가능성을 촉진합니다. 프리 라딕. Biol. 메드. 2020, 160, 111–124. [상호참조]
9. 스리바스타바, A.; 리카리, D.; 프라단, B.; 바라드와즈, KK; Gaballo, A.; 콰르타, A.; 예나, M.; 스리바스타바, S.; Ragusa, A. 대장암 생물학의 신경펩티드 억제제에 대한 통찰: 기회 및 중개 관점. 신청 과학. 2022, 12, 8990. [상호참조]
10. 구티에레스-로드리게스, AG; 후아레스-포르티야, C.; 올리바레스-바뉴엘로스, T.; Zepeda, RC 해조류의 항암작용. 약물 발견. 오늘 2018, 23, 434–447. [상호참조]
11. 지트, BP; 패트나익, S.; 아리아, R.; 대시, R.; 사후, SS; 프라단, B.; 부얀, PP; 베헤라, PK; 예나, M.; 샤르마, A.; 외. 식물화학물질(Phytochemicals): 방사선 보호를 위한 잠재적인 차세대 물질입니다. Phytomed. 국제 J. Phytother. 파이토팜. 2022, 2022, 154188. [CrossRef] [PubMed]
12. 지트, BP; 프라단, B.; 대시, R.; 부얀, PP; 베헤라, C.; 베헤라, RK; 샤르마, A.; 알카라즈, M.; Jena, M. Phytochemicals: 방사선 유발 신호 경로의 잠재적인 치료 조절제. 항산화제 2022, 11, 49. [CrossRef] [PubMed]
13. 파니그라히, GK; 야다브, A.; 만달, P.; 트리파티, A.; Das, M. Cassia occidentalis 씨앗의 안트라퀴논 부분인 라인의 면역조절 잠재력. 독성. 레트 사람. 2016, 245, 15–23. [상호참조] [PubMed]
14. 오바제, P.; 로마, A.; 스테클, M.; 니콜레티, L.; 아르나손, JT; Pandey, S. 주류 암 치료제로서 자연 건강 제품의 연구 및 개발을 발전시킵니다. 분명하다. 기반 보완. 대체. 메드. 2015, 2015, 751348. [CrossRef] [PubMed]
15. 파트라, S.; 나약, R.; 패트로, S.; 프라단, B.; 사후, B.; 베헤라, C.; 부티아, SK; Jena, M. 식이성 식물화학물질의 화학적 다양성과 화학적 예방 방식. 생명공학. Rep.(Amst. Neth.) 2021, 30, e00633. [상호참조]
16. 파트라, S.; 프라단, B.; 나약, R.; 베헤라, C.; 다스, S.; 파트라, SK; 에퍼스, T.; 예나, M.; Bhutia, SK 암의 화학 예방 및 시너지 효과에 대한 식이 폴리페놀: 임상 증거 및 분자 작용 메커니즘. Phytomed. 국제 J. Phytother. 파이토팜. 2021, 90, 153554. [CrossRef] [PubMed]
17. 파트라, S.; 프라단, B.; 나약, R.; 베헤라, C.; 판다, KC; 다스, S.; Jena, M. 암 치료제에서 식이성 식물화학물질을 조절하는 세포사멸 및 자가포식: 현재 증거 및 미래 전망. 식물원. 결의안. 2021, 35, 4194-4214. [상호참조]
18. 파트라, S.; 프라단, B.; 나약, R.; 베헤라, C.; 루트, L.; 예나, M.; 에퍼스, T.; Bhutia, SK 암에 대한 커큐민 및 레스베라트롤의 화학요법 효능: 화학 예방, 화학 보호, 약물 시너지 효과 및 임상 약동학. 암 생물학 세미나 진행 중; 학술 출판물: 미국 매사추세츠주 케임브리지, 2021; 310~320쪽.
19. 윌슨, RM; Danishefsky, SJ 치료제 발견에 있어서 소분자 천연물: 합성 연결. J.Org. 화학. 2006, 71, 8329-8351. [상호참조] [PubMed]
20. 시몬스, TL; 안드리아나솔로, E.; 맥파일, K.; 플랫, P.; Gerwick, WH 해양 천연물을 항암제로 사용합니다. 몰. 암 거기. 2005, 4, 333-342. [상호참조]
21. 아칸소주 캐롤; 콥, BR; 데이비스, RA; 키저스, RA; Prinsep, MR 해양 천연 제품. Nat. 찌르다. 2019, 36, 122–173. [상호참조] [PubMed]
22. Shinde, P.; 바네르지, P.; Mandhare, A. 신약의 원천으로서의 해양 천연 제품: 특허 검토(2015-2018). 전문가 의견. 거기. 가볍게 두드리기. 2019, 29, 283–309. [상호참조] 23. Pradhan, B.; 김현수; 압바시, S.; 기, J.-S. 해양 식물성 플랑크톤 독소의 독성 효과 및 종양 촉진 활동: 검토. 독소 2022, 14, 397. [CrossRef]
24. 프라단, B.; 나약, R.; 부얀, PP; 파트라, S.; 베헤라, C.; 사후, S.; Ki, J.-S.; 콰르타, A.; 라구사, A.; Jena, M. 항산화 조절에 관한 새로운 항균제로서의 M. Algal 플로로탄닌: 현재 발전과 미래 방향. 3월 Drugs 2022, 20, 403. [CrossRef] [PubMed]
25. 프라단, B.; 기, J.-S. 식물성 플랑크톤 독소와 잠재적인 치료 응용: 강력한 의약품을 향한 여정. 3월 Drugs 2022, 20, 271. [CrossRef] [PubMed]
26. 프라단, B.; 마하라나, S.; 박타, S.; Jena, M. 규조류와 와편모충류에 대한 새로운 기록을 특별히 언급한 인도 오디샤 해안의 해양 식물성 플랑크톤 다양성. 야채 2021, 35, 330–344. [상호참조]
27. 베헤라, C.; 대시, SR; 프라단, B.; 예나, M.; Adhikary, SP 인도 오디샤 안수파 호수의 조류 다양성. Nelumbo 2020, 62, 207–220. [상호참조]
28. 베헤라, C.; 프라단, B.; 팬더, R.; 나약, R.; 나약, S.; Jena, 인도, Huma(Ganjam), Saltpans의 M. Algal 다양성. J. 인디언 봇. Soc. 2021, 101, 107–120. [상호참조]
29. 대시, S.; 프라단, B.; 베헤라, C.; Jena, M. Kanjiahata 호수의 조류 다양성, Nandankanan, Odisha, 인도. J. 인디언 봇. Soc. 2020, 99, 11–24. [상호참조]
30. 대시, S.; 프라단, B.; 베헤라, C.; 나약, R.; Jena, 인도 오디샤 주 차트라푸르의 탬파라 호수에 있는 M. Algal Flora. J. 인디언 봇. Soc. 2021, 101, 1–15. [상호참조]
31. 마하라나, S.; 프라단, B.; 예나, M.; Misra, MK 인도 오디샤 Chilika 석호에 있는 식물성 플랑크톤의 다양성. 환경. Ecol 2019, 37, 737-746.
32. 모한티, S.; 프라단, B.; 파트라, S.; 베헤라, C.; 나약, R.; Jena, M. 해안 오디샤(Odisha)에서 분리된 일부 조류 계통의 영양 생리활성 화합물 스크리닝. J. 고급. 식물 과학. 2020, 10, 1–8.
33. 프라단, B.; 파트라, S.; 대시, SR; 사타파시, Y.; 나약, S.; 만달, 알래스카; Jena, M. 인도 오디샤의 Chilika 석호에서 수집한 해양 조류 Enteromorpha 압축의 시험관 내 항당뇨병, 항염증 및 항균 활성. 야채 2022, 35, 614–621. [상호참조]
34. 프라단, B.; 나약, R.; 파트라, S.; 부얀, PP; 베헤라, PK; 만달, 알래스카; 베헤라, C.; Ki, J.-S.; 아디카리, SP; 무바라크 알리, D.; 외. 바이러스 감염을 퇴치하는 항바이러스제로서의 후코이단에 대한 최신 리뷰입니다. 탄수화물. 폴리엠. 2022, 2022, 119551. [CrossRef] [PubMed]
35. 프라단, B.; 나약, R.; 파트라, S.; 부얀, PP; 대시, SR; Ki, J.-S.; 아디카리, SP; 라구사, A.; Jena, M. Cyanobacteria 및 항바이러스제로서의 조류 유래 생리활성 대사산물: 증거, 작용 방식 및 추가 확장 범위; SARS-CoV-2 발생에 따른 종합적인 검토. 항산화제 2022, 11, 354. [CrossRef] [PubMed]
36. 프라단, B.; 파트라, S.; 대시, SR; 나약, R.; 베헤라, C.; Jena, M. 항산화 조절에 대한 특별한 언급을 통해 Chlorella vulgaris Beyerinck [Beijerinck]의 메탄올 추출물의 항균 활성 평가. 퓨처제이팜. 과학. 2021, 7, 17. [상호참조]
37. 프라단, B.; 파트라, S.; 베헤라, C.; 나약, R.; 지트, BP; Ragusa, A. Enteromorpha intestina lis 추출물의 항산화, 항당뇨병 및 항염증 활성에 대한 예비 조사. Molecules 2021, 26, 1171. [CrossRef]
38. 프라단, B.; 파트라, S.; 베헤라, C.; 나약, R.; 파틸, S.; 부티아, SK; Jena, M. Enteromorpha 압축 추출물은 구강암에서 세포사멸과 자가포식을 통해 항암 활성을 유도합니다. 몰. Biol. 2020, 47, 9567-9578. [상호참조]
39. 쿠마시, A.; NA, 우샤코바; 메인 프레오브라젠스카야; D'Incecco, A.; 피콜리, A.; 토타니, L.; 티나리, N.; 모로제비치, GE; 버먼, AE; 미시간주 빌란 갈조류에서 추출한 9가지 후코이단의 항염증, 항응고, 항혈관신생 및 항부착 활성에 대한 비교 연구입니다. 당생물학 2007, 17, 541-552. [상호참조]
40. 박, HY; 한, MH; 박씨; 진, C.-Y.; 김지영; 최I.-W.; 김, 노다; 남, T.-J.; 권태경; Choi, YH 지질다당류로 유도된 BV2 미세아교세포에서 NF-κB, MAPK 및 Akt 활성화 억제를 통한 후코이단의 항염증 효과. 식품화학. 독성. 2011, 49, 1745-1752. [상호참조]